二恶英类化学物质毒性的分子机制还没完全研究清楚,但经过二十多年的研究人们对其机理也有了一定的认识。总的说来二恶英类化学物质产生作用并不是通过直接的损伤,二恶英类化学 物质并不与蛋白质和核酸形成加合物,也不直接损害细胞DNA一、芳香烃受体介导的基因表达
通过芳香烃受体介导基因表达(如P4501A1)是二恶英类化学物质毒性作用最主要也是最基本的作用机制。芳香烃受体是一高分子量的蛋白质(110-150KD),与二恶英类化学物质有可逆转的高亲和力,主要存在于细胞浆中(也有小部分在胞核中),其作用模式类似于甾体类受体,但也有不同。该蛋白属于basichelix-loop-helix PAS(Per-Arnt-Stim)超家族,该家族均为转录因子),均含有两个功能部位即:basichelix-loop-helix部位和PAS的复合物无活性的形式存在,除自身外还有3-4种蛋白质与之结合,其中只鉴别出了90 KD的热休克蛋白(heatshock protein, HSP90),该蛋白对受体的活性具有重要影响。
芳香烃受体介导的基因表达基本的作用过程可区分以下几个基本过程:①二恶英类化学物进入细胞;②配体-受体复合物与DNA识别位点结合;④特异基因的转录及翻译;⑤表达蛋白发挥作用。在这些过程中,前三步研究的较清楚,而后续过程还不是很清楚.
1. 2. 芳香烃复合物的形成并不能与DNA℃形成的复合物并不能正确与基因位点结合,而在高于在20℃形成的复合物则有生物活性,说明该过程需要温度依赖性的激活步骤。HSP90对受体的激活起着重要作用,HSP90结合(设想为抑制受体与核中的转录因子结合)。当配体与受体结合,原结合于受体的HSP90即脱落下来,暴露出受体的DNA 3. 受体复合物与DNA的结合。二恶英类化学物质与芳香烃受体的结合使芳香受体激活,随后配体-受体复合物即转移入胞,在细胞核中聚集。该复合物在与DNA结合以前必须与细胞核中的一种蛋白结合即芳香烃受体核转位子蛋白(Ah receptor nuclear translocator protein, ARNT结合的能力。该蛋白质分子量为87kDa超家族,含有两个功能部位即:basic helix-loop-helix(bHLH)部位和PAS的结合意义重大。仅保留芳香烃受体核转位子蛋白的bHLH部位,可保存ARNT形成二聚体及与DNA部位的两个α-helilx结构则仅与DNA部位包括两个亚结构即PASA和PAS B复合物然后与特异基因上游部位的增强子即二恶英反应元件(dioxin responsive element, XRE外来物反应元件的核心序列为5′-T/GNGCGTGA/CG/CA-3′ 4. 和1A2,谷胱甘肽S和1A2,同时也研究的最为广泛。AhR/ARNT复合物与增强子核心序列结合后,通过何种方式激活基因的转录研究较少。一般而言AhR/ ARNT复合物与增强子核心序列的结合后可导致DNA链的弯曲,核染色质的断裂,从而增加了激活启动子的机率,增加了CYP1A1起始转录的机率,导致细胞色素P4501A1在核中的聚集。Roberton等研究发现,在细胞色素P4501A1转录起始点上游,281-950个碱基间有九个顺式反应元件,其中三个为二恶英反应元件,另六个元件的作用不详。但当AhR/ ARNT方式.即进入细胞浆,结合于核糖体开始蛋白质的翻译。
5. 和1A2表达产物的研究,如:芳烃羟化酶,可将前致癌物转化为致癌物,从而促进机体癌症的发生。
以上为二恶英类化学物质介导机体基因表达的基本方式,目前对该过程的研究仍是二恶英类化学物导致毒性的主要研究方向。
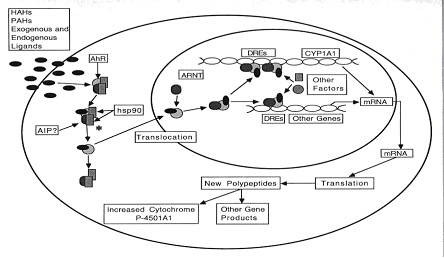
Fig.二、芳香烃受体介导的蛋白激酶途径
|